
Finally. I finally get round to doing the final part of the Tyrannosaurus rex pages. This one will look at how the animal lived (or rather, how we think it lived).


Erickson, G., Lappin, A. and Larson, P. (2005) 'Androgynous rex - The utility of chevrons for determining the sex of crocodilians and non-avian dinosaurs', Zoology, 108 (4), pp. 277-286
Erickson, G., Currie, P., Inouye, B. and Winn, A. (2006) 'Tyrannosaur life tables: an example of nonavian dinosaur population biology', Science, 313 (5784), pp. 213-217, doi: 10.1126/science.1125721
Horner, J. and Lessem, D. (1993) The complete T. rex, New York City: Simon and Schuster
Horner, J. and Padian, K. (2004) 'Age and growth dynamics of Tyrannosaurus rex', Proceedings of the Royal Society B, 271 (1551), pp. 1875-1880, doi: 10.1098/rspb.2004.2829
Hutchinson, J., Bates, K., Molnar, J., Allen, V. and Mackovicky, P. (2011) 'A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny and Growth,' PLoS One, 6 (10), e26037, doi: 10.1371/journal.pone.0026037
Kaye, T., Gaugler, G. and Sawlowicz, Z. (2008) 'Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms', PLoS One, 3 (7): e2808, doi: 10.1371/journal.pone.0002808
Lambe, L. (1914) 'On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon', Ottawa Naturalist, 27, pp. 129-135
Larson, P. (1994) 'Tyrannosaurus sex', in Rosenberg, G. and Wolberg, D. (eds) Dino Fest. The Paleontological Society Special Publications, 7, pp. 139-155
Lee, A., and Werner, S. (2008) 'Sexual maturity in growing dinosaurs does not fit reptilian growth models', Proceedings of the National Academy of Sciences, 105 (2), pp. 582-587, doi: 10.1073/pnas.0708903105
Leidy, J. (1858) 'On the bones of a huge herbivorous saurian near Haddonfield', Proceedings of the Academy of Natural Sciences of Philadelphia, 10, pp. 215-218
Leidy, J. (1865) 'Memoir on the extinct reptiles of the Cretaceous formations of the United States', Smithsonian Contributions to Knowledge, 14, pp. 1-135
Newman, B. (1970) 'Stance and gait in the flesh-eating Tyrannosaurus', Biological Journal of the Linnean Society B, 2 (2), pp. 119-123, doi: 10.1111/j.1095-8312.1970.tb01707.x
Osborn, H. (1905) 'Tyrannosaurus and other Cretaceous carnivorous dinosaurs', Bulletin of the American Museum of Natural History, 21 (14), pp. 259-265
Osborn, H. (1917) 'Skeletal adaptations of Ornitholestes, Struthiomimus and Tyrannosaurus', Bulletin of the American Museum of Natural History, 35 (43), pp. 733-771
Osborn, H. and Brown, B. (1906) 'Tyrannosaurus, Upper Cretaceous carnivorous dinosaur', Bulletin of the American Museum of Natural History, 22 (16), pp. 281-296
Paul, G. (1988) 'The Extreme Life Style and Habits of the Gigantic Tyrannosaurid Superpredators of the Cretaceous North America and Asia', in Larson, P. and Carpenter K. (eds) Tyrannosaurus, the Tyrant King, pp. 307-345, Bloomington: Indiana University Press
Peterson, J., Lenczewski, M. and Scherer, R. (2010) 'Influence of Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil and Extant Archosaurs', PLoS One, 5 (10), e13334, doi: 10.1371/journal.pone.0013334
Ross, R., Duggan-Haas, D. and Allmon, W. (2013) 'The Posture of Tyrannosaurus rex: Why Do Student Views Lag Behind the Science?', Journal of Geoscience Education, 61, p. 145, doi: 10.5408/11-259.1
Life History
Because of the number of fossils of Tyrannosaurus rex discovered, palaeontologists have a pretty good idea of the life of the dinosaur. From the fossils we have been able to see that T. rex probably lived into its late 20s (Erickson et al, 2004). We also know that T. rex experienced a major growth spurt during its life, growing from 1.8 tons at 14 to 6 tons at 18. Major teenage munchies then. After this period of major growth, the rate plateaus until old age. For example, only 600 kg separates 28-year-old Sue (the largest T. rex specimen discovered to date), a 22-year-old specimen discovered in Canada (Erickson et al., 2004). Other studies have confirmed this but some seem to suggest this plateauing occurred at a younger age, around 16 (Horner and Padian, 2004). Another study found that some individuals of Tyrannosaurus rex grew even faster, putting on an average 1800 kgs a year (Hutchinson et al., 2011). That's heavier than an average car! This sudden growth is likely to be due to physical maturity, as evidenced by a 16-20 year-old specimen discovered with medullary tissue in the femur, dubbed "B-rex". Medullary tissue is found only in female birds when ovulating (producing eggs), suggesting that B-rex was sexually mature (Schweitzer et al., 2005a). Further study has indicated that B-rex is 18-years-old (Lee and Werning, 2008). Other tyrannosaurs experienced a similar growth curve, however, because of their smaller adult size, their growth rates were smaller (Erickson et al., 2006). Over half of the Tyrannosaurus rex fossils found have been fully mature adults. This may be because juveniles had a low mortality rate, an incomplete fossil record or the excellent reason that fossil collectors are more interested in big, impressive bones rather than piddly little baby ones (Erickson et al., 2006). Or maybe it's because adults liked to live dangerously (Paul, 1988).
Sexual Dimorphism
Because of the sheer number of Tyrannosaurus rex fossils found, scientists have noticed that there appears to be 2 types or morphs, a robust morph and a gracile morph. Further analysis suggests that the robust morphs were female as suggested by the possession of wider hips (Carpenter, 1992) and smaller chevrons (pointy bits on the underside of the tail bones) (Larson, 1994), both interpreted as being adaptations for egg-laying.
Sounds like everything's pretty nailed down right? If only. The "females have smaller chevrons" idea was based on modern crocodiles and this was fine until someone discovered that not all female crocodiles have small chevrons. In fact, there was no correlation between chevron size and gender. This puts the Tyrannosaurus rex implication in doubt (Erickson et al., 2005). The most famous Tyrannosaurus fossil, "Sue" has turned out to have actually possessed a whacking great big chevron, despite being of the robust morph (Brochu, 2003).
So, back to the drawing board then? Not quite. Remember that B-rex specimen mentioned earlier? It possessed medullary tissue, which is used by modern female birds as a source of calcium for producing eggshells. Maybe that is a good indicator of gender? Unfortunately, B-rex is, so far, the only dinosaur know to have such tissue. But maybe if we compare B-rex with other specimens we might find other indicators?

Sounds like everything's pretty nailed down right? If only. The "females have smaller chevrons" idea was based on modern crocodiles and this was fine until someone discovered that not all female crocodiles have small chevrons. In fact, there was no correlation between chevron size and gender. This puts the Tyrannosaurus rex implication in doubt (Erickson et al., 2005). The most famous Tyrannosaurus fossil, "Sue" has turned out to have actually possessed a whacking great big chevron, despite being of the robust morph (Brochu, 2003).
So, back to the drawing board then? Not quite. Remember that B-rex specimen mentioned earlier? It possessed medullary tissue, which is used by modern female birds as a source of calcium for producing eggshells. Maybe that is a good indicator of gender? Unfortunately, B-rex is, so far, the only dinosaur know to have such tissue. But maybe if we compare B-rex with other specimens we might find other indicators?
Posture
Traditionally, Tyrannosaurus rex was depicted as standing upright, with the tail held low, kind of like a kangaroo. This idea originated from Joseph Leidy's reconstruction of the hadrosaur Hadrosaurus (Leidy, 1858, 1865). From then until the 1970s, most bipedal dinosaurs were reconstructed in this way. However, by the 1970s, palaeontologists realised that this was infeasible due to the excellent reason that this would have dislocated the hips and skull (Newman, 1970). However, this inaccuracy persisted in popular culture and the media until the release of Jurassic Park in 1993 (Ross et al., 2013).

Arms
Tyrannosaurus rex is well known for possessing very short arms. But why? That's a very good question. Originally, when it was first discovered. the only bone known was the humerus (Osborn, 1905). When the first skeletal reconstruction was shown to the public in 1915, Osborn based the forearms on the long, three-fingered ones present in the unrelated Allosaurus (Osborn, 1917). However, a year earlier, Lawrence Lambe described the short forearm of Tyrannosaurus' close relative Gorgosaurus (Lambe, 1914). This led to the belief that Tyrannosaurus had short forearms too, but this was not proven until the discovery of the specimen dubbed "Wankel rex" in 1989 (Horner and Lessem, 1993). "Sue" also contained complete forelimbs (Brochu, 2003). The first explanation given by Osborn, is that the arms were used as graspers during mating (Osborn and Brown, 1906). Maybe they were used to help lift the animal up from a prone position (Newman, 1970).
The most likely possibility, however, is that the arms were used to grab hold of struggling prey whilst the jaws did the business. This is based on the huge muscles present in the arms, 3 and a half times large than ours. The bones were also thickened to add additional strength. However, they were not very flexible, being limited to 40-45 degrees at the elbow, in contrast to a raptor's 88-130 degrees or a human's 165 (Carpenter and Smith, 2001).
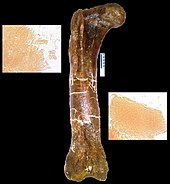
The most likely possibility, however, is that the arms were used to grab hold of struggling prey whilst the jaws did the business. This is based on the huge muscles present in the arms, 3 and a half times large than ours. The bones were also thickened to add additional strength. However, they were not very flexible, being limited to 40-45 degrees at the elbow, in contrast to a raptor's 88-130 degrees or a human's 165 (Carpenter and Smith, 2001).
Soft tissue
In 2005, the discovery of Tyrannosaurus rex bones with soft tissue preserved was announced. Blood vessels, bone tissue and even blood cells were found, with very similar structures to modern-day ostriches. However, it is currently unknown if any DNA is present so let's not start any Jurassic Park rumours yet (Schweitzer et al., 2005b).
Another big find in 2007 found, not just preserved tissue, but actual proteins. This is huge, as beforehand, palaeontologists presumed that fossils of this age have their proteins replaced with minerals. If more material of this nature is found, it could lead to the new field of palaeobiochemistry (Asara et al., 2007).
But, since 2008, there has been a debate over the veracity of the finds, beginning with Thomas Kaye and colleagues who argued that the proteins were actually the remnants of bacterial residues produced during fossilisation (Kaye, et al., 2008). Both sides have argued back and forth, with the last word being from Joseph Peterson and colleagues who demonstrated that they couldn't be from bacteria (Peterson et al., 2010).
I could sit here and talk about Tyrannosaurus for hours, but I think this will do. That wraps up the Tyrannosaurus series.
See also:
More dinosaurs
Tyrannosaurus rex Part 1
Another big find in 2007 found, not just preserved tissue, but actual proteins. This is huge, as beforehand, palaeontologists presumed that fossils of this age have their proteins replaced with minerals. If more material of this nature is found, it could lead to the new field of palaeobiochemistry (Asara et al., 2007).
But, since 2008, there has been a debate over the veracity of the finds, beginning with Thomas Kaye and colleagues who argued that the proteins were actually the remnants of bacterial residues produced during fossilisation (Kaye, et al., 2008). Both sides have argued back and forth, with the last word being from Joseph Peterson and colleagues who demonstrated that they couldn't be from bacteria (Peterson et al., 2010).
I could sit here and talk about Tyrannosaurus for hours, but I think this will do. That wraps up the Tyrannosaurus series.
See also:
More dinosaurs
Tyrannosaurus rex Part 1
References
Asara, J., Schweitzer, M., Freimark, L., Phillips, M. and Cantley, L. (2007) 'Protein sequences from Mastodon and Tyrannosaurus rex, revealed by mass spectrometry', Science, 316 (5822), pp. 280-285, doi: 10.1126/science.1137614
Brochu, C. (2003) 'Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull', Society of Vertebrate Paleontology Memoirs, 7, pp. 1-138
Carpenter, K. (1992) 'Variation in Tyrannosaurus rex', in Carpenter, K. and Currie, P. (eds) Dinosaur Systematics: Approaches and Perspectives, pp. 141-145, Cambridge: Cambridge University Press
Carpenter, K. and Smith, M. (2001) 'Forelimb Osteology and Biomechanics of Tyrannosaurus rex', in Tanke, C. and
Erickson, G., Makovicky, P., Currie, P., Norell, M., Yerby, S. and Brochu, C. (2004) 'Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs', Nature, 430 (7001), pp. 772-775, doi: 10.1038/nature02699
Asara, J., Schweitzer, M., Freimark, L., Phillips, M. and Cantley, L. (2007) 'Protein sequences from Mastodon and Tyrannosaurus rex, revealed by mass spectrometry', Science, 316 (5822), pp. 280-285, doi: 10.1126/science.1137614
Brochu, C. (2003) 'Osteology of Tyrannosaurus rex: insights from a nearly complete skeleton and high-resolution computed tomographic analysis of the skull', Society of Vertebrate Paleontology Memoirs, 7, pp. 1-138
Carpenter, K. (1992) 'Variation in Tyrannosaurus rex', in Carpenter, K. and Currie, P. (eds) Dinosaur Systematics: Approaches and Perspectives, pp. 141-145, Cambridge: Cambridge University Press
Carpenter, K. and Smith, M. (2001) 'Forelimb Osteology and Biomechanics of Tyrannosaurus rex', in Tanke, C. and
Erickson, G., Makovicky, P., Currie, P., Norell, M., Yerby, S. and Brochu, C. (2004) 'Gigantism and comparative life-history parameters of tyrannosaurid dinosaurs', Nature, 430 (7001), pp. 772-775, doi: 10.1038/nature02699
Erickson, G., Lappin, A. and Larson, P. (2005) 'Androgynous rex - The utility of chevrons for determining the sex of crocodilians and non-avian dinosaurs', Zoology, 108 (4), pp. 277-286
Erickson, G., Currie, P., Inouye, B. and Winn, A. (2006) 'Tyrannosaur life tables: an example of nonavian dinosaur population biology', Science, 313 (5784), pp. 213-217, doi: 10.1126/science.1125721
Horner, J. and Lessem, D. (1993) The complete T. rex, New York City: Simon and Schuster
Horner, J. and Padian, K. (2004) 'Age and growth dynamics of Tyrannosaurus rex', Proceedings of the Royal Society B, 271 (1551), pp. 1875-1880, doi: 10.1098/rspb.2004.2829
Hutchinson, J., Bates, K., Molnar, J., Allen, V. and Mackovicky, P. (2011) 'A Computational Analysis of Limb and Body Dimensions in Tyrannosaurus rex with Implications for Locomotion, Ontogeny and Growth,' PLoS One, 6 (10), e26037, doi: 10.1371/journal.pone.0026037
Kaye, T., Gaugler, G. and Sawlowicz, Z. (2008) 'Dinosaurian Soft Tissues Interpreted as Bacterial Biofilms', PLoS One, 3 (7): e2808, doi: 10.1371/journal.pone.0002808
Lambe, L. (1914) 'On a new genus and species of carnivorous dinosaur from the Belly River Formation of Alberta, with a description of the skull of Stephanosaurus marginatus from the same horizon', Ottawa Naturalist, 27, pp. 129-135
Larson, P. (1994) 'Tyrannosaurus sex', in Rosenberg, G. and Wolberg, D. (eds) Dino Fest. The Paleontological Society Special Publications, 7, pp. 139-155
Lee, A., and Werner, S. (2008) 'Sexual maturity in growing dinosaurs does not fit reptilian growth models', Proceedings of the National Academy of Sciences, 105 (2), pp. 582-587, doi: 10.1073/pnas.0708903105
Leidy, J. (1858) 'On the bones of a huge herbivorous saurian near Haddonfield', Proceedings of the Academy of Natural Sciences of Philadelphia, 10, pp. 215-218
Leidy, J. (1865) 'Memoir on the extinct reptiles of the Cretaceous formations of the United States', Smithsonian Contributions to Knowledge, 14, pp. 1-135
Newman, B. (1970) 'Stance and gait in the flesh-eating Tyrannosaurus', Biological Journal of the Linnean Society B, 2 (2), pp. 119-123, doi: 10.1111/j.1095-8312.1970.tb01707.x
Osborn, H. (1905) 'Tyrannosaurus and other Cretaceous carnivorous dinosaurs', Bulletin of the American Museum of Natural History, 21 (14), pp. 259-265
Osborn, H. (1917) 'Skeletal adaptations of Ornitholestes, Struthiomimus and Tyrannosaurus', Bulletin of the American Museum of Natural History, 35 (43), pp. 733-771
Osborn, H. and Brown, B. (1906) 'Tyrannosaurus, Upper Cretaceous carnivorous dinosaur', Bulletin of the American Museum of Natural History, 22 (16), pp. 281-296
Paul, G. (1988) 'The Extreme Life Style and Habits of the Gigantic Tyrannosaurid Superpredators of the Cretaceous North America and Asia', in Larson, P. and Carpenter K. (eds) Tyrannosaurus, the Tyrant King, pp. 307-345, Bloomington: Indiana University Press
Peterson, J., Lenczewski, M. and Scherer, R. (2010) 'Influence of Microbial Biofilms on the Preservation of Primary Soft Tissue in Fossil and Extant Archosaurs', PLoS One, 5 (10), e13334, doi: 10.1371/journal.pone.0013334
Ross, R., Duggan-Haas, D. and Allmon, W. (2013) 'The Posture of Tyrannosaurus rex: Why Do Student Views Lag Behind the Science?', Journal of Geoscience Education, 61, p. 145, doi: 10.5408/11-259.1
Schweitzer, M., Wittmeyer, J. and Horner, J. (2005a) 'Gender-specific reproductive tissue in ratites and Tyrannosaurus rex', Science, 308 (5727), pp. 1456-1460, doi: 10.1126/science.1112158
Schweitzer, M., Wittmeyer, J., Horner, J. and Toporski, J. (2005b) 'Soft-tissue vessels and cellular preservation in Tyrannosaurus rex', Science, 307 (5717), pp. 1952-1955, doi: 10.1126/science.1108397
Schweitzer, M., Wittmeyer, J., Horner, J. and Toporski, J. (2005b) 'Soft-tissue vessels and cellular preservation in Tyrannosaurus rex', Science, 307 (5717), pp. 1952-1955, doi: 10.1126/science.1108397